MOVIMIENTO NUCLEAR INTERKINETICO Y NOTCH DURANTE LA NEUROGENESIS
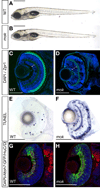
Las neuronas surgen durante el desarrollo de un epitelio pseudoestratificado columnar.Los diferentes tipos celulares en el SNC derivan de un pool común de progenitores. Los núcleos de estos progenitores se mueven entre el lado apical y basal del neuroepitelio, según la fase del ciclo celular en la que se encuentren, proceso que se denomina Migración Nuclear Interkinética (INM).La retina neural, consiste en 6 tipos neuronales y gliales, los que se diferencian según el orden de su nacimiento. En las retinas de mutantes de Zf mikre oko ( mok) , en los cuales la proteína dinactina-1 está disfuncional, la INM es más rápida y más profunda hacia el lado basal y más lenta hacia el apical del neuroepitelio. Los mutantes de Zebrafish mok, presentan una retina cuyos progenitores dejan el ciclo celular prematuramente, acelerándose la neurogénesis y produciéndose un desequilibrio entre las células nuevas y antiguas.Paralelamente se estudio cómo influía este cambio en el movimiento en la señalización de Notch, el cual se activa en el lado apical. En Drosophila, Notch, a través de la inhibición lateral, mantiene la vecindad celular en un estado proliferativo multipotente, mientras que la disminución de esta señal es un pre-requisito para que se produzca la diferenciación neuronal.Los datos obtenidos en el estudio demuestran que la función del INM es balancear la exposición a señales proliferativas tanto neurogénicas como proliferativas de los precursores del neuroepitelio.
RESULTADOS
1) MUTANTES MOK TIENEN EXCESO DE CÉLULAS GANGLIONARES EN LA RETINA
Como en otros mutantes, Zf mok carecen de fotorreceptores a los 5 dpf, aunque nacen con ellos y van muriendo por apoptosis entre el día 2.5 y 3 dpf.Presentan también un aumento de la capa de células ganglionares ( GCL) comparado con wild type. Así mismo se encuentran células ganglionares en la capa nuclear interna (gran número de células progenitoras se vuelven competentes para producir CGR).
2) CÉLULAS BIPOLARES O DE LA GLIA DE MÜLLER ESTÁN DISMINUIDAS O AUSENTES
En la retina, todos los tipos neuronales y as células gliales de Müller, derivan de un progenitor común. Se especula entonces que la superproducción de CGR llevaría a una depleción del pool progenitor. Para investigar esto se estudió la presencia de marcadores para otros tipos celulares.Se concluyó que la glía de Müller está disminuída o ausente en retinas de Zf mok a los 5dpf ( Inmunohistoquímica para glutamina sintetasa). Lo mismo ocurrió para células bipolares ( disminución de PKCalfa)
3) GRAN NÚMERO DE PROGENITORES DEJAN EL CICLO CELULAR EN ZF MOK
Los factores que retardan el fin del ciclo celular en la retina, dan lugar a un exceso de células post mitóticas, incrementando la generación de CGR.Para investigar el timing de estas células, se marcaron con IdU y BrdU entre las 28 y 36 hpf respectivamente. Los mutantes mok mostraron un aumento de IdU y no señal del BrdU, evidenciando las células dejaban el ciclo celular antes , durante la primera oleada de la neurogénesis.
4) EL GEN MOK CODIFICA DINACTINA-1
Se clonó posicionalmente el gen mok. Su fenotipo presenta una mutación puntual en la Dinactina-1Por WB, realizado en extractos de embriones enteros, se vio que la Dnct-1 se detecta en wt y en zf mok en los 3 primeros días, indicando que son suplidos maternalmente. Desapareciendo la proteína a los 4 dpf.
5) INM ESTÁ PERTURBADO EN MUTANTES MOK
En Drosophila, ensayos de pérdida de función de la dinactina-1, revelaron que es indispensable para una correcta migración celular y posicionamiento del núcleo.Se especula que en los mutantes mok, el INM está alterado y eso tendría su efecto en el término del ciclo celular.El resultado obtenido fue una migración rápida y más profunda hacia la superficie basal, llevando más tiempo el retorno y entrando en mitosis antes de alcanzar la superficie apical.
6) GRADIENTE DE NOTCH CREA DISTINTOS GRADOS DE SEÑALIZACIÓN A LO LARGO DEL EJE APICO-BASAL
La activación de Notch endentece la neurogénesis. ARN de Notch1a, predomina en la región apical del neuroepitelio de la retina en Zf. Los ligandos del receptor Notch, DeltaB y DeltaC, se ubican además en la mitad basal de la retina.Después de unirse a su ligando, el receptor Notch produce un clivaje proteolítico. El fragmento citosólico se trasloca al núcleo donde modula la transcripción de varios genes.Se plantea un gradiente de Notch a lo largo del eje ápico-basal.
7) INM DIRIGIDO POR DINACTINA SIRVE PARA EXPONER LOS PROGENITORES AL GRADIENTE DE NOTCH
La distribución diferencial de Notch, junto con el INM, regulan la tasa de neurogénesis. Cómo sucede esto? El núcleo moviéndose más basalmente regula en baja a Notch, mientras que los núcleos que se mantienen en la luz ventricular retienen altos niveles de actividad Notch, impidiendo su diferenciación neurogénica a ese nivel.
CONCLUSIONES
- La mutación de la proteína Dinactina-1 perturba el INM y produce una superproducción de neuronas tempranas en la retina de los mutantes mok de Zf.
- El núcleo de los progenitores migra más rápido y más profundo en dirección basal y más lento en dirección apical.
- Notch ( señal antineurogénica) es más rica en la región apical en wt y mutantes mok.
- Notch previene a los progenitores de dejar el ciclo celular y diferenciarse prematuramente. La inhibición de Notch, produce aspectos similares al mutante mok.
- Entre mitosis , los núcleos de los progenitores se mueven hacia el gradiente Notch. Si el núcleo permanece cercano al polo apical, encontrará niveles de Notch elevados, y se quedará como célula proliferativa. Por otro lado, si el núcleo se trasloca más basalmente, Notch estará reducido, predisponiendo al progenitor a producir hijas neuronas durante las subsecuentes mitosis.
- INM y la señalización Notch, son puntos comunes a lo largo del neuroepitelio del SNC, así como en los epitelios no neurales, siendo este mecanismo usado en gran medida no sólo en el desarrollo, sino en el crecimiento y la regeneración.
0 comentarios