ENCUENTRO DEL PROGRAMA ACORTANDO DISTANCIAS


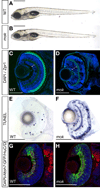
Foto del Taller 2011. Maestras observando un embrión de pez cebra -coloreado por técnicas inmunocitoquímicas- en el microscopio confocal del Instituto Clemente Estable
ANEP - PEDECIBA
Talleres en temas científico-tecnológicos para Maestros:
El Taller de Postgrado para Maestros
Vuelve en 2012 en dos Locaciones:
Investigador Responsable: Dr. Daniel Rodríguez-Ithurralde, Laboratorio de Neurociencia Molecular, Instituto Clemente Estable (IIBCE).
Breve descripción del taller
Se busca que los maestros experimenten directamente las interrogantes, planteamientos, formulación de hipótesis, experimentación y análisis crítico inherentes a la investigación científica.
La actividad teórica y los talleres de discusión serán los necesarios para justificar y planificar la actividad práctica en el laboratorio. El desarrollo del pez cebra (Danio rerio) será documentado en fotos, videos y preparados microscópicos, con énfasis en los procesos de desarrollo. Además, se ilustrará el uso biotecnológico de los peces para el monitoreo ambiental y en ensayos de toxicidad de sustancias.
Se espera que quienes culminen la Parte 2 demuestren no solamente habilidad para analizar críticamente las imágenes y datos experimentales obtenidos, sino también la capacidad de producir nuevos preparaciones y materiales docentes, a emplear en sus clases o en las de sus pares. Eventualmente, alguno de los maestros querrá seguir el camino de Estable.
Información e Inscripciones on line:
El enlace con la página del Consejo de Enseñanza Inicial y Primaria (CEIP), donde se encuentra el anuncio del Taller y el formulario de Inscripción en línea, es el siguiente:
http://www.cep.edu.uy/index.php?option=com_content&view=article&id=1513&Itemid=853
Por consultas: Al Departamento de Formación en Servicio, ANEP-CEIP,
Tel/Fax 2916 6307, Cel. 091 413441
Al Director del Taller, IIBCE, Dr. Daniel Rodríguez-Ithurralde: Tel. 24871616

| Corydoras paleatus nacido en nuestro laboratorio (R) | ||
Oganizados por el
Laboratorio de Neurociencia Molecular
Instituto de Investigaciones Biológicas Clemente
Estable (IIBCE)
Lugar: Sala Uppsala,
Instituto de Investigaciones Biológicas Clemente Estable,
Av. Italia 3318, Montevideo
Hora: 16:30
Ciclo: Receptores Sensoriales de los Peces y su Desarrollo
- - Lic. Gabriela del Puerto. - Lunes 27 de febrero
- - Dr. Daniel Rodríguez-Ithurralde.- Lunes 12 de marzo.
- - Prof. Alicia Soza. - Martes 27 de marzo.
- - Maestra Andrea Etchartea. - Martes 10 de abril.
- Lic. G. del Puerto Vitovich. Martes 15 de mayo.
- Dr. Daniel Rodríguez-Ithurralde. Martes 29 de mayo.
- Prof. Andrea Echartea. Martes 12 de junio.
- Lic . Gabriela del Puerto. Martes 26 de junio.
La asistencia a los Seminarios es gratuita pero se requiere inscripción previa (ver abajo). Se entregará Diploma de Asistencia
Inscripción: Las personas interesadas en concurrir a los Seminarios deberán inscribirse, enviando al Coordinador de los mismos, Dr. Daniel Rodríguez-Ithurralde (Email: drit@iibce.edu.uy o drithurralde@gmail.com), un E-mail que incluya nombre, documento de identidad, dirección de e-mail y teléfono del interesado, donde se explique en una frase la motivación o interés en asistir. Si lo desea y es el caso, puede incluir, lugar de trabajo, aunque no es imprescindible.
How Chemicals Affect Us, By NICHOLAS D. KRISTOF Published: May 2, 2012
Scientists are observing with increasing alarm that some very common hormone-mimicking chemicals can have grotesque effects. A widely used herbicide acts as a female hormone and feminizes male animals in the wild. Thus male frogs can have female organs, and some male fish actually produce eggs. In a Florida lake contaminated by these chemicals, male alligators have tiny penises. These days there is also growing evidence linking this class of chemicals to problems in humans. These include breast cancer, infertility, low sperm counts, genital deformities, early menstruation and even diabetes and obesity. Philip Landrigan, a professor of pediatrics at Mount Sinai School of Medicine, says that a congenital defect called hypospadias — a misplacement of the urethra — is now twice as common among newborn boys as it used to be. He suspects endocrine disruptors, so called because they can wreak havoc with the endocrine system that governs hormones.
Endocrine disruptors are everywhere. They’re in thermal receipts that come out of gas pumps and A.T.M.’s. They’re in canned foods, cosmetics, plastics and food packaging. Test your blood or urine, and you’ll surely find them there, as well as in human breast milk and in cord blood of newborn babies. In this campaign year, we are bound to hear endless complaints about excessive government regulation. But here’s an area where scientists are increasingly critical of our government for its failure to tackle Big Chem and regulate endocrine disruptors adequately. Last month, the Endocrine Society, the leading association of hormone experts, scolded the Food and Drug Administration for its failure to ban bisphenol-A, a common endocrine disruptor known as BPA, from food packaging.
Last year, eight medical organizations representing genetics, gynecology, urology and other fields made a joint call in Science magazine for tighter regulation of endocrine disruptors. Shouldn’t our government be as vigilant about threats in our grocery stores as in the mountains of Afghanistan? Researchers warn that endocrine disruptors can trigger hormonal changes in the body that may not show up for decades. One called DES, a synthetic form of estrogen, was once routinely given to pregnant women to prevent miscarriage or morning sickness, and it did little harm to the women themselves. But it turned out to cause vaginal cancer and breast cancer decades later in their daughters, so it is now banned. Scientists have long known the tiniest variations in hormone levels influence fetal development. For example, a female twin is very slightly masculinized if the other twin is a male, because she is exposed to some of his hormones. Studies have found that these female twins, on average, end up slightly more aggressive and sensation-seeking as adults but have lower rates of eating disorders. Now experts worry that endocrine disruptors have similar effects, acting as hormones and swamping the delicate balance for fetuses in particular. The latest initiative by scholars is a landmark 78-page analysis to be published next month in Endocrine Reviews, the leading publication in the field. “Fundamental changes in chemical testing and safety determination are needed to protect human health,” the analysis declares. Linda S. Birnbaum, the nation’s chief environmental scientist and toxicologist, endorsed the findings. The article was written by a 12-member panel that spent three years reviewing the evidence. It concluded that the nation’s safety system for endocrine disruptors is broken. “For several well-studied endocrine disruptors, I think it is fair to say that we have enough data to conclude that these chemicals are not safe for human populations,” said Laura Vandenberg, a Tufts University developmental biologist who was the lead writer for the panel. Worrying new research on the long-term effects of these chemicals is constantly being published.
One study found that pregnant women who have higher levels of a common endocrine disruptor, PFOA, are three times as likely to have daughters who grow up to be overweight. Yet PFOA is unavoidable. It is in everything from microwave popcorn bags to carpet-cleaning solutions. Big Chem says all this is sensationalist science. So far, it has blocked strict regulation in the United States, even as Europe and Canada have adopted tighter controls on endocrine disruptors. Yes, there are uncertainties. But the scientists who know endocrine disruptors best overwhelmingly are already taking steps to protect their families. John Peterson Myers, chief scientist at Environmental Health Sciences and a co-author of the new analysis, said that his family had stopped buying canned food. “We don’t microwave in plastic,” he added. “We don’t use pesticides in our house. I refuse receipts whenever I can. My default request at the A.T.M., known to my bank, is ‘no receipt.’ I never ask for a receipt from a gas station.” I’m taking my cue from the experts, and I wish the Obama administration would as well.
• I invite you to visit my blog, On the Ground. Please also join me on Facebook and Google+, watch my YouTube videos and follow me on Twitter.

Autores: Andrea Etchartea, Alicia Soza, Isabella Benvenuto, Gabriela del Puerto Mónica González Lario, Verónica Gargiulo, Zully Bornia, Daniel Rodríguez-Ithurralde[1]
Taller desarrollado en el Laboratorio de Neurociencia Molecular del Instituto de Investigaciones Biológicas Clemente Estable (IIBCE).
PRESENTACIÓN
El estudio del desarrollo embrionario de vertebrados nos ofrece importantes indicios de cómo se va generando la organización anatómica y funcional del ser adulto. Muchas veces nos permite, además, explicar cómo se producen algunas enfermedades o malforma-ciones.
¿POR QUÉ EL PEZ CEBRA?
El pez cebra (Danio rerio), banderita o zebrafish (ZF) es muy utilizado en investigación básica y aplicada, debido a que, entre muchas otras virtudes, sus embriones son transparentes y se desarrollan fuera del cuerpo de la madre, permitiendo seguir en el estereomicros-copio, las transformaciones morfoló-gicas y funcionales que ocurren en su cuerpo desde el primer minuto postfecundación.
PROCEDIMIENTOS EXPERIMENTALES
Se emplearon los distintos aumentos de un estereomicroscopio para observar y describir las características morfológicas y fisiológicas normales de huevos, embriones y larvas vivas (en 3-D) y ver la respuesta a distintos estímulos (luz, temperatura, fármacos, etc.) en función del tiempo (4-D). Se reconocieron los distintos estadíos evolutivos, y se compararon con una tabla estándar (Kimmel, 1995). Se puso especial atención en la aparición y modificaciones de las placodas ópticas y óticas, a partir de las 24 hpf. Algunos de los ejemplares fueron anestesiados con hidrato de cloral y fijados con paraformaldehído al 4 % y otros fijadores, con el fin de preservar sus tejidos. El material fue luego sometido a técnicas histológicas que pusieron en evidencia, en forma diferencial, distintos componentes o células del organismo (técnicas de impregnación argéntica, de inmunofluorescencia, etc.) y observado con el microscopio óptico, confocal y/o electrónico de transmisión, de acuerdo a los procedimientos usuales en el IIBCE.
RESULTADOS OBTENIDOS:
I.- IDENTIFICACIÓN DE LAS PRINCIPALES ETAPAS DEL DESARROLLO
Desarrollo temprano
El desarrollo se inició con la segmentación (divisiones celulares sucesivas) del huevo fecundado, con un rápido incremento del número de células. Fue seguida por la gastrulación, caracterizada por movimientos morfogenéticos que reorganizan las células primero en dos y luego en tres capas: ectodermo, endodermo y mesodermo, dando origen a los distintos órganos y sistemas del adulto.
Neurulación
Es el proceso que conduce a la formación del tubo neural, que luego dará origen a las distintas regiones del sistema nervioso central. Esta fase no la observamos en preparados.
Embrión somítico
Se caracteriza por la segmentación del mesodermo en bloques llamados somites. A partir de estos somites se formará el esqueleto, músculos y dermis de las distintas regiones del cuerpo. Observamos esta etapa en la cual comienzan a desarrollarse los órganos internos de los distintos aparatos y se produce un importante desarrollo diferencial de distintas regiones del sistema nervioso. Este período se extiende hasta las 24 hpf.
Eclosión
Comprobamos que entre las 48 -72 se rompe la membrana coriónica y se libera la larva al medio circundante. Conservaron aún por unos días cierta cantidad de vitelo, con el cual se nutre los primeros cuatro días.
Período larvario
En este estadío continuó la formación de órganos primarios, los cartílagos de la cabeza y las aletas. Observamos los latidos cardíacos, la circulación sanguínea, los movimientos corporales y diversas respuestas a estímulos mecánicos.
Juvenil
Presentaron la mayoría de las características del adulto, pero en individuos de menor tamaño.
Adulto
Se considera adulto a partir de los tres meses, período en que alcanza la madurez sexual y comienzan a observarse con más claridad las diferencias morfológicas entre macho y hembra (dimorfismo sexual).
II.- Desarrollo de los sistemas sensoriales
Luego de abordar el desarrollo general de estos peces, profundizamos en su relacionamiento con el mundo circundante, usando como modelo a los sistemas sensoriales.
El universo sensorial de los peces es totalmente diferente al nuestro
“Los peces tienen excelentes sentidos de la vista y olfato y un exclusivo sistema de la línea lateral, que con exquisita sensibilidad detecta corrientes de agua y vibraciones proporcionando un tacto a distancia en el agua” (Hickman, 2009). A partir de las primeras 24 hpf observamos con el microscopio estereoscópico, la aparición y desarrollo de las placodas ópticas y óticas, las cuales dan lugar a la formación de los ojos y los oídos, respectivamente.
Una sensibilidad especial para vertebrados acuáticos.
La densidad del agua la hace un excelente conductor de vibraciones. No sorprende que los organismos acuáticos puedan detectar estas señales mecánicas (vibraciones) por distintas mecanismos de mecano-recepción, que aparecen muy temprano en la historia de los vertebrados y han sido altamente modificados y especializados en los peces (Helfman, 2009).
La mecanorecepción implica la detección de movimientos del agua donde vive el pez. Los peces tienen dos grandes sistemas mecanosensoriales: el sistema de la línea lateral y el oído interno. Ambos se basan en las células sensoriales pilosas las cuales contienen un penacho de prolongaciones parecidas a cilias en su superficie apical (Hickman, 2009; Kardong, 2006; Helfman, 2009).
Con técnicas inmunohistoquímicas logramos observar los trayectos neurales correspondientes a la inervación de los canales cefálicos y corporales del sistema de la línea lateral, tal como se ve coloreado de rojo en la figura.
CONCLUSIONES Y PERSPECTIVAS
Se realizó la observación, registro y fotografía de embriones, larvas y adultos del pez cebra a través de las distintas etapas del desarrollo.
En lo referente al desarrollo de los órganos sensoriales, visualizamos los esbozos de los mismos. La temprana aparición de las placodas óptica y óticas, y su importante tamaño en relación con otras estructuras, permitió comprender la importancia de estos órganos para tempranos requerimientos ambientales y para la supervivencia en la etapa larvaria, por ejemplo.
En lo que respecta al sentido de la línea lateral, tan característico de los peces, comprobamos la abundancia en la región cefálica de los receptores sensoriales denominados neuromastos, viendo su distribución en distintos territorios.
En cuanto a las repercusiones de esta instancia semi-presencial, podemos afirmar que permitió un crecimiento en lo disciplinar de los participantes, y la comprobación de la utilidad de la enseñanza basada en la evidencia, en lo pedagógico. Tuvimos la oportunidad de manejar diferentes recursos y hacerlo en grupo. El trabajo en taller permitió desarrollar nuevas estrategias, haciendo del intercambio de ideas y del trabajo conjunto una forma de enriquecimiento profesional.
El éxito de las jornadas nos obliga a plantearnos la necesidad de integrar experiencias como éstas en el desarrollo del currículo.
“Todo el mundo parece estar de acuerdo que para los jóvenes de hoy,
es importante saber algo del mundo de la ciencia y que es tarea de la escuela
garantizar que esto ocurra.”
Guy Claxon 1
Bibliografía
Claxton, G. (1991). Educar mentes curiosas. Ed. Visor: Madrid.
Helfman, G. (2009). The diversity of fishes. Wiley-Blackwell: London.
Hickman, F. (2009). Principios generales de zoología. 13ª Edición. McGraw-Hill: Barcelona.
Kimmel, Ch. and Ballard, W. (1995). The stages of embryonic development of the zebrafish. Developmental Dynamics, 203: 253-310.
[1] Dirigir toda correspondencia a Dr. Daniel Rodríguez-Ithurralde, Lab. de Neurociencia Molecular, IIBCE - Av. Italia 3318, Montevideo – E-mail: drit@iibce.edu.uy – httm://www.iibce.edu.uy/NMOLF/index.html
Las neuronas surgen durante el desarrollo de un epitelio pseudoestratificado columnar.Los diferentes tipos celulares en el SNC derivan de un pool común de progenitores. Los núcleos de estos progenitores se mueven entre el lado apical y basal del neuroepitelio, según la fase del ciclo celular en la que se encuentren, proceso que se denomina Migración Nuclear Interkinética (INM).La retina neural, consiste en 6 tipos neuronales y gliales, los que se diferencian según el orden de su nacimiento. En las retinas de mutantes de Zf mikre oko ( mok) , en los cuales la proteína dinactina-1 está disfuncional, la INM es más rápida y más profunda hacia el lado basal y más lenta hacia el apical del neuroepitelio. Los mutantes de Zebrafish mok, presentan una retina cuyos progenitores dejan el ciclo celular prematuramente, acelerándose la neurogénesis y produciéndose un desequilibrio entre las células nuevas y antiguas.Paralelamente se estudio cómo influía este cambio en el movimiento en la señalización de Notch, el cual se activa en el lado apical. En Drosophila, Notch, a través de la inhibición lateral, mantiene la vecindad celular en un estado proliferativo multipotente, mientras que la disminución de esta señal es un pre-requisito para que se produzca la diferenciación neuronal.Los datos obtenidos en el estudio demuestran que la función del INM es balancear la exposición a señales proliferativas tanto neurogénicas como proliferativas de los precursores del neuroepitelio.
RESULTADOS
1) MUTANTES MOK TIENEN EXCESO DE CÉLULAS GANGLIONARES EN LA RETINA
Como en otros mutantes, Zf mok carecen de fotorreceptores a los 5 dpf, aunque nacen con ellos y van muriendo por apoptosis entre el día 2.5 y 3 dpf.Presentan también un aumento de la capa de células ganglionares ( GCL) comparado con wild type. Así mismo se encuentran células ganglionares en la capa nuclear interna (gran número de células progenitoras se vuelven competentes para producir CGR).
2) CÉLULAS BIPOLARES O DE LA GLIA DE MÜLLER ESTÁN DISMINUIDAS O AUSENTES
En la retina, todos los tipos neuronales y as células gliales de Müller, derivan de un progenitor común. Se especula entonces que la superproducción de CGR llevaría a una depleción del pool progenitor. Para investigar esto se estudió la presencia de marcadores para otros tipos celulares.Se concluyó que la glía de Müller está disminuída o ausente en retinas de Zf mok a los 5dpf ( Inmunohistoquímica para glutamina sintetasa). Lo mismo ocurrió para células bipolares ( disminución de PKCalfa)
3) GRAN NÚMERO DE PROGENITORES DEJAN EL CICLO CELULAR EN ZF MOK
Los factores que retardan el fin del ciclo celular en la retina, dan lugar a un exceso de células post mitóticas, incrementando la generación de CGR.Para investigar el timing de estas células, se marcaron con IdU y BrdU entre las 28 y 36 hpf respectivamente. Los mutantes mok mostraron un aumento de IdU y no señal del BrdU, evidenciando las células dejaban el ciclo celular antes , durante la primera oleada de la neurogénesis.
4) EL GEN MOK CODIFICA DINACTINA-1
Se clonó posicionalmente el gen mok. Su fenotipo presenta una mutación puntual en la Dinactina-1Por WB, realizado en extractos de embriones enteros, se vio que la Dnct-1 se detecta en wt y en zf mok en los 3 primeros días, indicando que son suplidos maternalmente. Desapareciendo la proteína a los 4 dpf.
5) INM ESTÁ PERTURBADO EN MUTANTES MOK
En Drosophila, ensayos de pérdida de función de la dinactina-1, revelaron que es indispensable para una correcta migración celular y posicionamiento del núcleo.Se especula que en los mutantes mok, el INM está alterado y eso tendría su efecto en el término del ciclo celular.El resultado obtenido fue una migración rápida y más profunda hacia la superficie basal, llevando más tiempo el retorno y entrando en mitosis antes de alcanzar la superficie apical.
6) GRADIENTE DE NOTCH CREA DISTINTOS GRADOS DE SEÑALIZACIÓN A LO LARGO DEL EJE APICO-BASAL
La activación de Notch endentece la neurogénesis. ARN de Notch1a, predomina en la región apical del neuroepitelio de la retina en Zf. Los ligandos del receptor Notch, DeltaB y DeltaC, se ubican además en la mitad basal de la retina.Después de unirse a su ligando, el receptor Notch produce un clivaje proteolítico. El fragmento citosólico se trasloca al núcleo donde modula la transcripción de varios genes.Se plantea un gradiente de Notch a lo largo del eje ápico-basal.
7) INM DIRIGIDO POR DINACTINA SIRVE PARA EXPONER LOS PROGENITORES AL GRADIENTE DE NOTCH
La distribución diferencial de Notch, junto con el INM, regulan la tasa de neurogénesis. Cómo sucede esto? El núcleo moviéndose más basalmente regula en baja a Notch, mientras que los núcleos que se mantienen en la luz ventricular retienen altos niveles de actividad Notch, impidiendo su diferenciación neurogénica a ese nivel.
CONCLUSIONES
- La mutación de la proteína Dinactina-1 perturba el INM y produce una superproducción de neuronas tempranas en la retina de los mutantes mok de Zf.
- El núcleo de los progenitores migra más rápido y más profundo en dirección basal y más lento en dirección apical.
- Notch ( señal antineurogénica) es más rica en la región apical en wt y mutantes mok.
- Notch previene a los progenitores de dejar el ciclo celular y diferenciarse prematuramente. La inhibición de Notch, produce aspectos similares al mutante mok.
- Entre mitosis , los núcleos de los progenitores se mueven hacia el gradiente Notch. Si el núcleo permanece cercano al polo apical, encontrará niveles de Notch elevados, y se quedará como célula proliferativa. Por otro lado, si el núcleo se trasloca más basalmente, Notch estará reducido, predisponiendo al progenitor a producir hijas neuronas durante las subsecuentes mitosis.
- INM y la señalización Notch, son puntos comunes a lo largo del neuroepitelio del SNC, así como en los epitelios no neurales, siendo este mecanismo usado en gran medida no sólo en el desarrollo, sino en el crecimiento y la regeneración.



Laboratorio de Neurociencia Molecular y Núcleo de Biotecnología y Marcadores Moleculares, Instituto Clemente Estable, Av. Italia 3318, Montevideo, Uruguay. E-mail: drit@iibce.edu.uy
Introduction
A growing body of evidence suggests that herbicides formulated in base of N-(phosphonomethyl)glycine (glyphosate), the most fequently used in the world, present considerable toxicity for algae, molluscs, fishes and amphybia (Quassinti et al., 2008). Its non-controlled use represents a real menace for biodiversity, natural fishery resources, and pisciculture.
It is of great interest to point out that non-permitted concentrations of this and other herbicides and/or their metabolites and/or coadjuvants have been found in waters and foods for human use, and these findings are a cause of concern as potential dangers for human and animal health, which must be strictly monitored. The most abundant glyphosate metabolite, the AMPA, is more toxic and more resistant to degradation than it. The aim of this work is to investigate and, when possible, quantify the effects of glyphosate on zebrafish (Danio rerio) development using internationally accepted tests (Hill et al, 2005; Baunbeck & Lammar, 2006).
Materials and Methods
Animals used. Zebrafish embryo produced in our facility according to internationally accepted procedures were used. We have applied our previously validated, multi-parametric, Zebrafish Embryo Toxicity Assay (ZETA), which allows measuring numerous morphologic, physiologic and biochemical parameters in hundreds of simultaneous samples with sensitivity, high reproductibility and low cost (Ojeda et al., 2007; Rodriguez-Ihurralde., 2007) to quantify glyphosate effects on zebrafish development.
ZETA test. Specimens of one post-fecundation hour of life (hpf) were incubated in ELISA plates at 27.5 ± 1ºC, at a one-per-well ratio. Each well contained 1.0 ml of filtered, controlled aquarium water containing a pre-established final concentraciones (ccf) of glyphosate. After fixed, pre-stablished time intervals (12, 24, 48, 72 and 96 hpf), the presence of universally acepted final points of zebrafish development was computed (Baunbeck & Lammar, 2006; Ojeda et al., 2007; Bortagaray et al., 2009).
Biochemical assays. All biochemical determinations were run in triplicate. Total cholinesterase, acetylcholinesterase (AChE) and butyrylcholinesterase (BChE) assays were performed as described in Karlsson et al. (1984), using acetylthiocholine as a substrate.
Results
Lethality. Lethality plots showed that from ccf of 25 microg/ml and higher, embryo lethality was significantly dependent on both the herbicide concentration and the time of exposure. At 24 hpf, lethalities of 35.4% were verified for ccf of 50 microg/ml, whereas it was 74% at ccf of 75 microg/ml, and 82% at ccf of 150 microg/ml, wheras in control embryos remained between 0-5%.
Ecclosion events. Although most morphologic-structural final points exhibited delays in their time of appearance in embryos exposed to glyphosate, ecclosion events were, as a general rule, accelerated. At ccf between 12.5-50 microg/ml, corion effraction with embryo ecclosion appeared earlier, i.e., the ratio of eclosioned/total embryos at 72 hpf was significanly higher than in controls. Therefore, arrive in direct contact with the external aquatic medium, forms which are more immature than in control samples.
Cardiac and pericardic alterations. Embryionic cardiac frequency, as measured at 48 hpf, decreased significatintly, from ccf 25 microg/ml on, with regard to control specimens without glyphosate. At ccf of 50 microg/ml, cardiac and pericardic edema, and delays in the separation of the viteline sac, appear.
Physiopatologic and biochemical changes. At ccf of 75 microg/ml or higher, all the embryos showed at least one physiologic or structural developmental alteration, as, for instance, cardio-pericardic edema, circulatory delay, cholinesterase inhibition, or decreases in trunk flection movements at stadii in which they should be present.
Digestivo-viteline malformations. Morphologic alterations of the viteline sac, or its absence of separation, are frequently seen. Some of the morphologic alteratins are so important, that were incompatible with life.
Musculo-eskeletic malformations. Deformities of varied importance were frequent in both axial and apendicular skeleton. Fishes showed incurvations, “gibas”, and angles of the vertebral axis and varied deformities and abnormal divisions of tail and appendages.
Discussion and Conclusions
In summary, we have verified a group of changes in parameters of type functional, biochemical and morphologic. There is a correlation between high concentrations and more severe alterations encountered, with the exeption of the cases where the toxicity is so high that the embryos are unable to survive more than one minutes or hours.
Early ecclosion might also be viewed as an additional cause of lethality, since the corion barrier, which is normally protecting immature embryos fom the external medium, is opened earlier in herbicide-treated specimens (Ojeda et al., 2007). This early release of immature forms to the water, when occurs in the usual environment of real water bodies, might cause massive deaths of embryos under the effect of other biological, physical or chemical agents, that can enter in direct contact with the embryo (Ojeda et al., 2007; Rodríguez-Ithurralde et al., 2007).
It is relevant to point out that we have demonstrated lethal and sub-lethal toxic effects, as well as teratogenic effects at glyphosate ranks of concentration which have been previously reported as atoxic. The concentrations used in our experiments are 1-3 orders of magnitude inferior to those determined in water sheds close to culturs in Argentina and Uruguay. Thus, in very exact measurements carried out in Pergamino, Argentina, in water bodies close to cultures treated with glyphosate, the levels found in the water varied along the year and along the day only between 0.1 and 0.7 mg/ml, i.e., they never reached levels under 0.1 mg/ml.
Acknowledgements
We are indebted to PEDECIBA (Biología) for its continued grant support to DRI and for fellowship support to AS.
References
Bortagaray, V., Barrios, L., Cruces Aramburu, R., Del Puerto, G., Ojeda, P., Saravia, A. Rodríguez-Ithurralde, D. (2009). Los ensayos de embriotoxicidad en pez cebra detectan una variedad de efectos teratogénicos del glifosato.
6th SBBM Jorn. Biotechnology Section, (Uruguay)
Braunbeck, T. & Lammar, E. (2006). Fish embryo toxicity assays. German Federal Environmental Agency, 1/298-40/298.
Ellman G. L., Courtney, K. D., Andres, V. & Featherstone, R. M., 1961. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol., 7, 88-95.
Hill, A.J., Teraoka, H., Heideman, W. and Peterson, R.E., 2005. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 86, 6-19.
Karlsson, E., Mbugua, P. & Rodríguez-Ithurralde, D., 1984. Fasciculins, anticholinesterase toxins from the venom of the green mamba Dendroaspis angusticeps. J. Physiol. (Paris), 79, 232-240.
Ojeda, M.P., Parnizari, F. & Rodríguez Ithurralde, D., 2007. Aspectos cuantitiativos del desarrollo embrionario del pez cebra (Danio rerio) y su validación para ensayos de toxicidad ambiental. Actas Fisiol. (Montevideo), 11: 52-52.
Olivera, S., Rodríguez-Ithurralde, D. & Henley, J. M., 2003. Acetylcholinesterase promotes neurite elongation, synapse formation and surface expression of AMPA receptors in hippocampal neurones. Mol. Cell. Neurosci., 23, 96-106.
Quassinti, L., Maccari, E., Murri, O. & Bramucci, M., 2008. Effects of paraquat and glyphosate on steroidogenesis in gonads of the frog Rana sculenta in vitro.
Rodríguez-Ithurralde, D. (2007). Biomarcadores en peces autóctonos y exóticos para el monitoreo eco-toxicológico de los recursos acuáticos. Actas Fisiol. (Montevideo), 11: 53-53.

Ier Congreso Uruguayo de Zoología, Facultad de Ciencias de Montevideo
5-10 de Diciembre 2010
Organizador y Moderador del Simposio: Dr. Daniel Rodríguez-Ithurralde
Laboratorio de Neurociencia Molecular y Toxicología del Desarrollo,
Instituto de Investigaciones Biológicas Clemente Estable (IIBCE).
Av. Italia 3318. E-mail: drit@iibce.edu.uy y drithurralde@gmail.com
PROGRAMA
■ Dr. Daniel Rodríguez-Ithurralde. Destaque introductorio: La espectacular diversidad sensorial de la inmensa radiación evolutiva de los peces.
■ Dr. Ángel Caputi. Diferenciación de las señales auto- y alo-generadas.
■ Dr. Pedro Aguilera. Discriminación de objetos de acuerdo a su impedancia en el pez de descarga débil Gymnotus omarorum
■ Dr. Daniel Rodríguez-Ithurralde y Andrea Saravia. Organización anatómica, neural y molecular del oído interno de los peces.
■ Dra. María E. Castelló. Desarrollo postnatal del sistema electrosensorial: papel de la proliferación celular en el crecimiento relativo de la estructuras neurales centrales.
■ Dra. Anabel Fernández. Proliferación y características celulares del tectum óptico y el torus longitudinalis de peces del género Austrolebias (Cyprinodontiformes – Rivulidae)
□ Discusión General y Conclusiones Finales

Durante el desarrollo desde un cigoto unicelular a un organismo maduro, numerosas divisiones celulares y fenómenos de diferenciación tienen lugar. La diferenciación y la formación de los tejidos necesitan estar coordinados con el ciclo celular para garantizar una correcta ocupación espacio-temporal en el plan corporal.La gastrulación que es el proceso temprano del desarrollo que lleva a la formación de 3 capas embrionarias (ecto, meso y endodermo) involucra toda una serie de movimientos celulares. En el zebrafish, la gastrulación comienza con la internalización de los precursores del mesodermo y endodermo en la región dorsal del embrión, en el anillo marginal. Existen varias teorías de cómo estas células penetran y comienzan a migrar debajo del epiblasto hacia el polo animal . Algunos sostienen que lo hacen individualmente por delaminación, y otros que las células superficiales alrededor del anillo lo hacen por involución.
Numerosos estudios destinados a investigar la coordinación entre todos estos procesos, proponen sistemas para modular el ciclo celular en los estadíos tempranos y observar luego cómo se comporta el embrión durante las etapas sucesivas de desarrollo. Entre estos modelos se encuentra el presentado en marzo 2009 por Xiaolin Liu y colaboradores, acceso en http://jcb.rupress.org/cgi/content/abstract/184/6/805 . Su modelo propone la manipulación del balance Geminina/Cdt1, produciendo cambios en el ciclo celular y por consiguiente también en los procesos apoptóticos que tienen lugar para modelar el embrión, produciendo también cambios en los niveles del Factor antiapoptótico nuclear kB (NF-kB). NF-kB se asocia con el promotor Snail1a y activa al represor transcripcional de su mismo nombre el cual juega un papel crucial en la delaminación y migración de las células del mesoendodermo, proponiendo un mecanismo molecular por el cual NF-kB y Snail1a se coordinan con el ciclo celular durante la gastrulación.
1) EL CICLO CELULAR ES MODULADO POR LA MANIPULACIÓN DEL BALANCE GEMININA / CDT1
Una vez que el ADN se comienza a replicar, el proceso debe ser inhibido durante el mismo ciclo celular para mantener la estabilidad genética del individuo. La Geminina se acumula en el núcleo desde la fase temprana S hasta el final de la mitosis y directamente se une a Cdt1, evitando que éste se una al MCM (complejo microcromosómico) previniendo así la resíntesis de ADN.La progresión en el ciclo celular es incompatible con ciertos tipos de conductas que se dan durante el desarrollo, como la migración, adhesión o cambios morfológicos. En su trabajo Xiaolin Liu y colaboradores ( marzo 2009) , establecieron un sistema para modelar el ciclo celular en la gastrulación manipulando el balance Geminina/Cdt1 en embriones de Zf.
Métodos: Aislamientos de cDNA para Geminina, Cdt1 y mrGem ( geminina resistente al MO) y mutagénesis de Geminina en diferentes aminoácidos.Morfolinos y m RNAs ( Geminina, Cdt1, MCM5, p65, p53, p21)
Resultados: En el estadío de blastodisco se observó una transcipción uniforme en los embriones de geminina, en tanto Cdt1 predominó en la zona dorsal, indicando que a ese nivel es mayor el número de mitosis. ( http://jcb.rupress.org/cgi/content/full/jcb.200806074/DC1) En los ejemplares inyectados con Gem MO, Cdt1 mRNA hubo un aumento de las mitosis, por el contrario en los que se produjo una sobre expresión de Gem o knockdown de Cdt1 , disminuyeron. Igual sucedió con los embriones inyectados con MCM5MO, se observó una disminución de las mitosis. Morfolino oligo contra p21, un inhibidor de las quinasas dependientes de ciclina mostró un aumento en las células proliferantes.
Todos estos resultados muestran que un adecuado balance entre geminina y Cdt1 es crítico para la regulación del ciclo celular, ya que la manipulación del mismo en más o en menos produce variaciones importantes en la gastrulación de los embriones.
2) LA MODULACIÓN DEL CICLO CELULAR PRODUCE DEFECTOS EN LOS MOVIMIENTOS CELULARES DE DELAMINACIÓN
Para agendar los efectos en la modulación del ciclo celular en la gastrulación del Zf, se observaron los fenotipos tratados con Gem MO en estadíos avanzados y se realizaron co-transplantes a aceptores wild type para indagar en profundidad los efectos de la modulación del ciclo en los movimientos mesoendodérmicos.
Métodos: Inyección de Gem MO y observación de fenotipos al final de la gastrulación. Expresión de Goosecoid (gsc) como marcador de la placa precordal. Ensayo de co-transplante de células de la placa mesodérmica precordal contro y con ciclo modulado a embriones aceptores en etapa de escudo de tipo salvaje.
Resultados: Se observa una disminución ántero-posterior del eje corporal con la inyección de Gem MO, lo que sugiere defectos en el movimiento del mesoendodermo. Para la expresión de gsc, se observó al 80% de la epibolia un retraso en la expresión cuando los ejemplares fueron inyectados con Gem MO y Cdt1mRNA.En el ensayo de co-transplante se dedujo que, el knockdown the Geminina llevaba a un retraso en la migración de las células y esto era revertido entre otras cosas por Snail1a.
Se concluyó entonces, que la delaminación que es el primer paso en la gastrulación y en los movimientos celulares durante la misma, está influenciada por las modulaciones del ciclo celular.
3) EL CICLO CELULAR COORDINA LOS MOVIMIENTOS CELULARES MESOENDODERMICOS A TRAVES DE LA APOPTOSIS, NF-Kb Y SU BLANCO DIRECTO, SNAIL1a.
La apoptosis juega un papel crucial en el moldeado del embrión durante la gastrulación, y está debidamente equilibrada con el ciclo celular. Se quiso investigar el mecanismo a través del cual el ciclo celular coordinaba los movimientos durante la gastrulación con la apoptosis en embriones con modulación de su ciclo.
Métodos: Ensayo TUNEL .Ensayo CAT. Ensayo ChIP
Resultados: Los niveles de apoptosis se correlacionan con el ciclo celular, y son dependientes de p53. P65 es un importante factor antiapoptótico en la gastrulación de los embriones de ZF.Al analizar el nivel nuclear de NF-kB se observó que es correlativo con el nivel de apoptosis en los embriones gastrulando.
La modulación del ciclo celular resulta en niveles anormales de la forma activa de NF-kB , y éste es el activador directo de Snail1a, lo que lleva a cambios en los movimientos celulares.
4) COORDINACION ENTRE EL CICLO CELULAR Y LOS MOVIMIENTOS MESOENDODÉRMICOS SE REQUIERE PARA UNA CORRECTA DISTRIBUCIÓN DE LOS PRECURSORES CELULARES
La gastrulación lleva a la formación de las 3 capas germinales. Después de la delaminación y la subsecuente migración , el hipoblasto contribuye a la configuración de dos capas: meso y endodermo.Defectos en la delaminación produce efectos en la formación de estas capas.Se observó la distribución del mesoendodermo con diferentes marcadores.
Métodos: Ensayo Casanova ( codifica una proteína relacionada con Sox). Expresión de no tail.
Resultados: Durante la gastrulación , el ciclo celular coordina la distribución de las células endodérmicas mediante apoptosis, NF-kB y Snail1a.
5) LOS DEFECTOS EN LA DISTRIBUCIÓN DE LAS CÉLULAS ENDODÉRMICAS CAUSADAS POR MODULACIÓN DEL CICLO CELULAR PRODUCE DEFECTOS EN LA INDUCCIÓN NEURAL.
La inducción neural está conectada al tejido endodérmico durante la gastrulación. Estudios previos reportaron que en Xenopus la sobreexpresión de geminina inducía al tejido neural a producirse de forma ectópica (Kroll et al.,1998). Se vio que la expresión de Otx2 y Sox2, estaba dispersa en embriones inyectados con Gem MO , lo que era parcialmente rescatada por la inyección de Snail1a mRNA. Para investigar si esta inducción neural atípica era secundaria a un defecto en la distribución del endodermo o no, se realizaron ensayos de transplante.
Método: Ensayo Casanova en embriones cas mutantes.
Resultado: Se recuperó la expresión de los marcadores neurales, tanto al 90% de la epibolia como hasta el estadío de 14 somites, aún en la ausencia de geminina en el ectodermo.
Esto indica que los defectos en la distribución de las células endodérmicas causados por la modulación en el ciclo lleva secundariamente a defectos en la inducción neural.
CONCLUSIONES
En el estudio que revisamos ( Xiaolin Liu et al,2009) se propone un sistema para manipular el ciclo celular en embriones tempranos de Zf, basado en el balance Geminina/Cdt1. Cambios estables en distintos niveles de la gastrulación ( escudo, 70%, 90% epibolia), indican la habilidad para manipular en positivo o negativo el ciclo celular de los embriones durante la gastrulación. Se concluye además que hay cambios recíprocos entre el número de apoptosis y el nivel de mitosis en las células activas. Estos cambios deberían variar luego de la gastrulación. Los resultados obtenidos muestran un aumento general en el estadío de 18 somites de la apoptosis en los ejemplares, ya sea inyectados con Gem MO o Cdt1MO, o geminina mRNA o Cdt1 mRNA. El autor plantea dos posibles razones para esto, ya sea que al modularse el ciclo y los movimientos gastrulatorios con su consiguiente variación de formación de las capas, aumenta el proceso apoptótico como regulador de la integridad general de los embriones. O que , alternativamente, la coordinación molecular entre el ciclo celular, la apoptosis y los movimientos celulares llegado un momento del desarrollo, cambia a otros mecanismos regulatorio, perdiendo coordinación. La apoptosis se observa solamente después de la gastrulación en la mayoría de los vertebrados. Sin embargo un nivel adecuado de la misma , junto con una maquinaria antiapoptótica que incluye en este caso a NF-kB y a Snail1a que es su blanco corriente abajo, es necesario para la coordinación entre el ciclo celular y los movimientos mesoendodermales. Se han hecho reportes previos de que Snail1a se expresa en la zona marginal que es la región de delaminación , y en la capa sincitial vitelínica (YSL), pero no en las células que mesoendodérmicas que migran. En este estudio se vio que Snail1a, es capaz autonómicamente de rescatar los movimientos defectuosos causados por la modulación del ciclo. Esto sugeriría que estas variaciones harían expresarlo en zonas que normalmente aparece.
En resumen, los investigadores proponen que la progresión en el ciclo celular , lo cual coincide con un nivel adecuado de apoptosis y de un contrabalance antiapoptótico ( el que involucra a NF-kB y a Snail1a) son críticos para la delaminación de las células del mesoendodermo y para la coordinación de sus movimientos durante la gastrulación.

Los peces como modelo de estudio en la biología de los vertebrados
Por Gaby Vitovich
Entre los organismos modelo se comparten mecanismos de regulación genética y de diferenciación celular que pueden ser estudiados y utilizados para entender mecanismos más complejos . Sin embargo, también pueden ser estudiados por tener características únicas.
Formalmente, “sistema modelo” u “organismo modelo” es la denominación que se aplica en biología, a las especies animales que se estudian en detalle y que se utilizan como base para generalizar como ocurren determinados procesos biológicos en el resto de las especies relacionadas. Pero, ¿existe realmente una especie modelo?. En realidad no. Depende del tipo de estudios que se quieran realizar y de hasta donde queramos generalizar los resultados obtenidos. Entre los distintos grupos de vertebrados candidatos a ser considerados “organismos modelo” para muchas de las disciplinas de la biología , los peces teleósteos (peces óseos) ocupan una de las primeras posiciones.
Filogenéticamente hablando, los peces constituyen el grupo de vertebrados mas antiguo (unos 500 millones de años) y diverso que se conoce. Agrupan aproximadamente unas 18.000 especies (un orden de magnitud mas que los mamíferos) y colonizan habitats muy diversos, por lo que han tenido que desarrollar estrategia evolutivas muy variadas. De estos tres rasgos evolutivos podemos ya señalar tres de las ventajas que poseen los peces como animales de experimentación:
1.-Ofrecen una gran cantidad de modelos alternativos. Su divergencia evolutiva y extrema diversidad proporciona una abundante fuente de genomas distintos donde buscar y encontrar todas las posibles combinaciones estructura-función que han ocurrido en la naturaleza durante los últimos 400 años .
2.-Son adecuados tanto para la experimentación de campo como la de laboratorio . Se adaptan fácilmente a la vida en cautividad y pueden mantenerse en los acuarios sin demasiados requerimientos técnicos.
3.-Su adquisición y mantenimiento son mucho más económicos que la de cualquier ave, anfibio, mamífero, etc, utilizado en experimentación .
Aunque con distinta finalidad, son muchas y variadas las especies de peces que se han utilizado y aún se utilizan en investigación, nos vamos a centrar en una especie perteneciente al grupo de los peces teleósteos,
El Pez cebra y la Biología de los vertebrados
En los últimos años se ha avanzado notablemente en la compresión del desarrollo y genética de los vertebrados y en gran medida el pez cebra es responsable de ello. Básicamente porque a partir de él, y con relativa facilidad, pueden generarse colecciones de mutantes cuyo fenotipo puede ser analizado iv vivo (en el propio animal) durante las etapas tempranas del desarrollo Alterar la función de un gen y observar el fenotipo que produce es una de las mejores alternativas para poder establecer cual es su función y que papel tiene en el desarrollo o funcionamiento de un organismo. Cuando tras inducir una mutación (mediante sustancias químicas], clonaje posicional, retrovirus genéticamente modificados, etc) el fenotipo mutante generado resulta de interés, se procede a localizar la mutación en el genoma y posteriormente a su clonaje. Mediante estas aproximaciones experimentales, se han desvelado importantes datos acerca de la formación y función de tejidos, órganos y redes neuronales e incluso mecanismos genéticos implicados en enfermedades humanas . Además, estos análisis en pez cebra están constituyendo un complemento esencial del “proyecto genoma humano” .
A la par de una descripción detallada de la anatomía del embrión del pez cebra se han desarrollado metodologías robustas que permiten la experimentación en varios aspectos como son: La localización de proteínas “in vivo” por medio de fusiones de la proteína verde fluorescente (GFP), el apagado específico de genes empleando morfolinos (RNA antisentido modificado con un grupo morfolino), la ablación y transplante de células.
¿Qué características posee el pez cebra frente a otros modelos que también permiten un sencillo análisis de genotipos mutantes (gusano y mosca de la fruta), para que a partir del se puedan generalizar datos sobre el desarrollo de vertebrados e incluso servir de modelo para el estudio de enfermedades humanas ?.
1.- Es un teleósteo diploide y su patrón de desarrollo es similar al de los vertebrados superiores incluidos los mamíferos y por lo tanto el hombre. El modelo Caenhorhabditis elegans / Drosophila deja muchas cuestiones por resolver porque la cresta neural, la notocorda, el páncreas endocrino y exocrino, etc, son “inventos” de los vertebrados. De hecho, en un principio “el sistema pez cebra” fue establecido como un puente para cubrir las lagunas existentes entre los modelos Drosophila/Caenhorhabditis elegans y ratón/hombre .
2.-Se ha demostrado la existencia relaciones sinténicas entre los genomas humanos y de pez cebra y por ello la mutación de genes ortólogos en pez cebra puede dar lugar a fenotipos similares a los que se presentan en muchas enfermedades humanas.
Frente al resto de los modelos de vertebrados posee como ventajas:
1.-Su pequeño tamaño (aproximadamente 2,5 cm) permite mantener un elevado número de ejemplares en espacios relativamente pequeños
2.-Una sola hembra madura puede poner desde docenas a cientos de huevos por lo que se puede disponer de elevadas cantidades de material genético uniforme
3.-La fertilización de los huevos es externa por lo que no se necesitan posteriores manipulaciones del embrión. De esta manera, se suprime la problemática que plante en mamíferos el tener que implantar posteriormente los embriones en el útero de hembras pseudopreñadas.
4.-Tanto el tiempo de desarrollo del embrión completo (72h) como el que transcurre entre distintas generaciones es muy corto (2-3 meses).Además, se pueden controlar las puestas modificando las temperatura y/o fotoperíodo.
5.-Los embriones son transparentes por lo que se permite el análisis no invasivo del embrión y el seguimiento la expresión génica en tiempo real (en el animal vivo según se desarrolla).
En la actualidad son incontables las investigaciones sobre biología del desarrollo, toxicología, carcinogénesis, senescencia, desordenes genéticos etc, que utilizan como modelo de estudio el pez cebra, y los logros obtenidos a partir de ellas imposibles de relatar. A modo de ejemplo vamos a citar la enfermedad de la Anemia Sideroblástica Congénita, en el que se refleja claramente el potencial del pez cebra como modelo de estudio de enfermedades humanas.
Mediante clonaje posicional se aislaron en zebrafish un conjunto de genes denominados Sauternes (Sau). Fenotípicamente, los mutantes Sau de zebrafish presentan bajos niveles de hemoglobina aunque el número de células rojas circulantes en el torrente sanguíneo es normal. Tras un intensivo estudio se demostró que este fenotipo mutante se debe a un defecto en el gen de la enzima d-aminolevulinato sintetasa (ALAS-2) que regula el primer paso en la biosíntesis del grupo hemo en los glóbulos rojos embrionarios. Los pacientes humanos de Anemia Sideroblástica Congénita también son deficientes en la enzima ALAS-2 y sus eritrocitos, al igual que los del pez cebra, hipocrómicos por deficiencias en la síntesis de hemoglobina. Para esta grave enfermedad, es la primera vez que se ha podido establecer un modelo animal de estudio.
Embriones de Zf con GFP

Zebrafish embryotoxicity in water quality monitoring.